
A Tropical Oasis Awaits!
Visit the Garden and explore the sights, sounds, and smells of the #1 Botanic Garden in the United States.
Purchase your tickets online today!

Jurassic Garden Returns!
Jurassic Garden returns! Embark on an awe-inspiring journey through time, surrounded by life-size dinosaurs nestled among our vast collection of cycads and other living plants from the Prehistoric World.





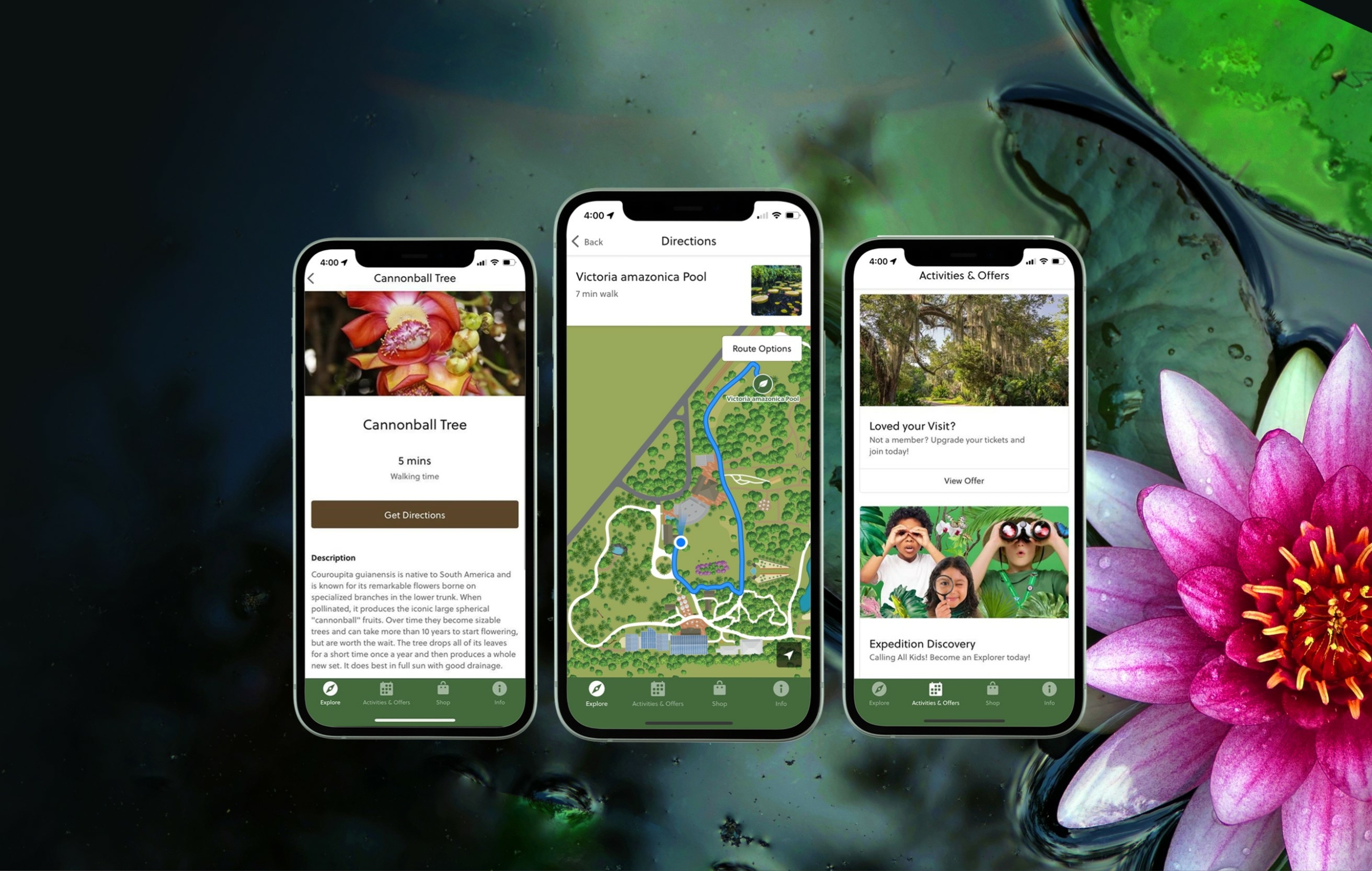






















Hours & Admission
Fairchild is open every day except for Christmas, December 25
Hours
10:00 a.m. – 5:00 p.m.
Admission
Fairchild Members Free
Adults $24.95
Child (6-17) $11.95
Students (with valid ID) $15.95
Seniors (65 and over) $17.95
In-Garden tram rides Free
Parking Free
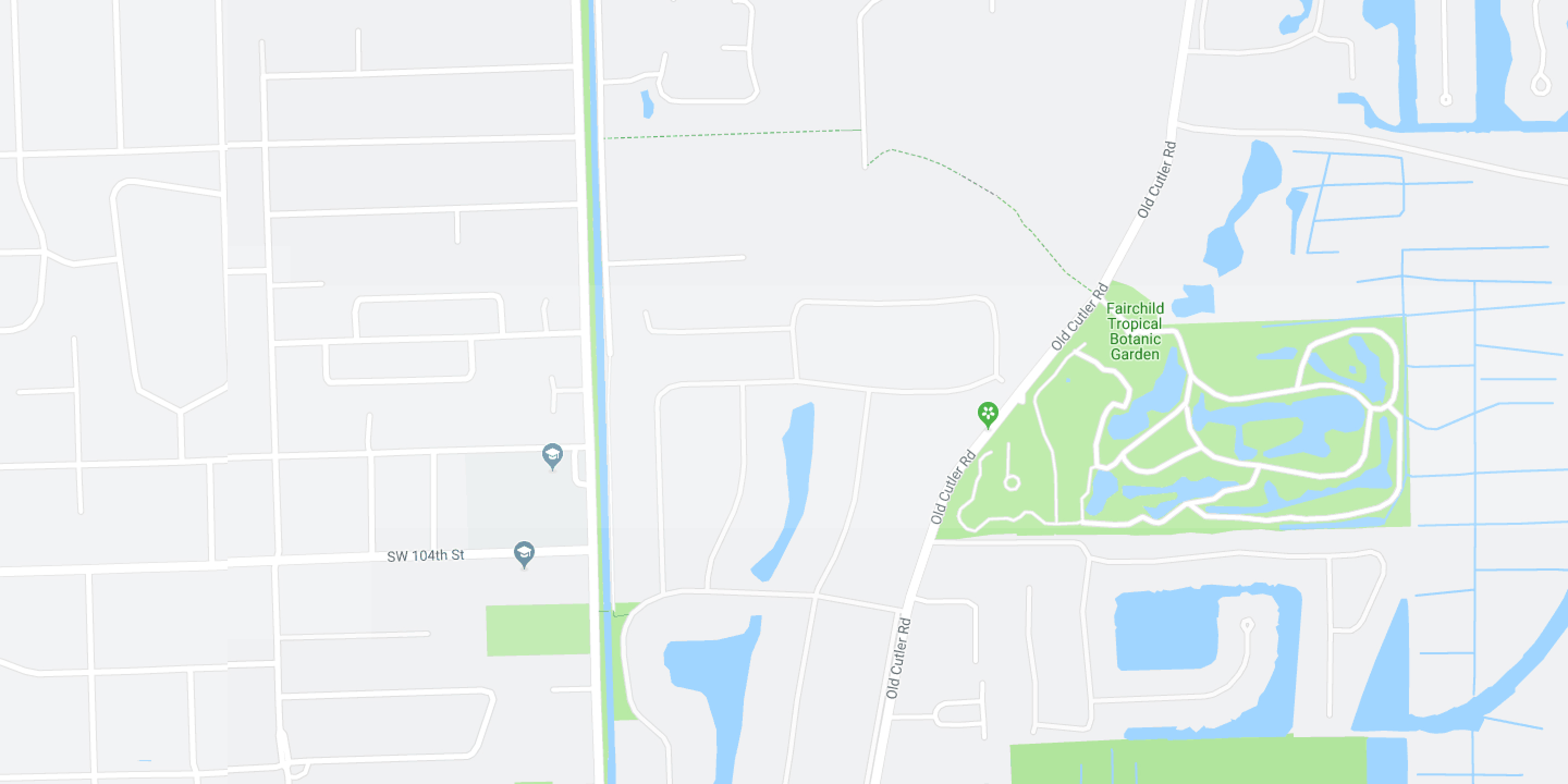
Location
10901 Old Cutler Rd, Miami, FL 33156
MembersGet Tickets Non-Members
Buy Tickets



FOLLOW THE LATEST @fairchildgarden