
Visit our Tropical Oasis
Explore the wonders and beauty of tropical plants and nature at Fairchild.

Garden of Shadows
Don’t miss two nights of Halloween fun at Fairchild Garden—wander spooky trails, sip festive cocktails, and uncover surprises at every turn!

Visit our Tropical Oasis
Explore the wonders and beauty of tropical plants and nature at Fairchild.

Orchids in October
Bring the whole family for a weekend of orchids, smiles, and October fun!






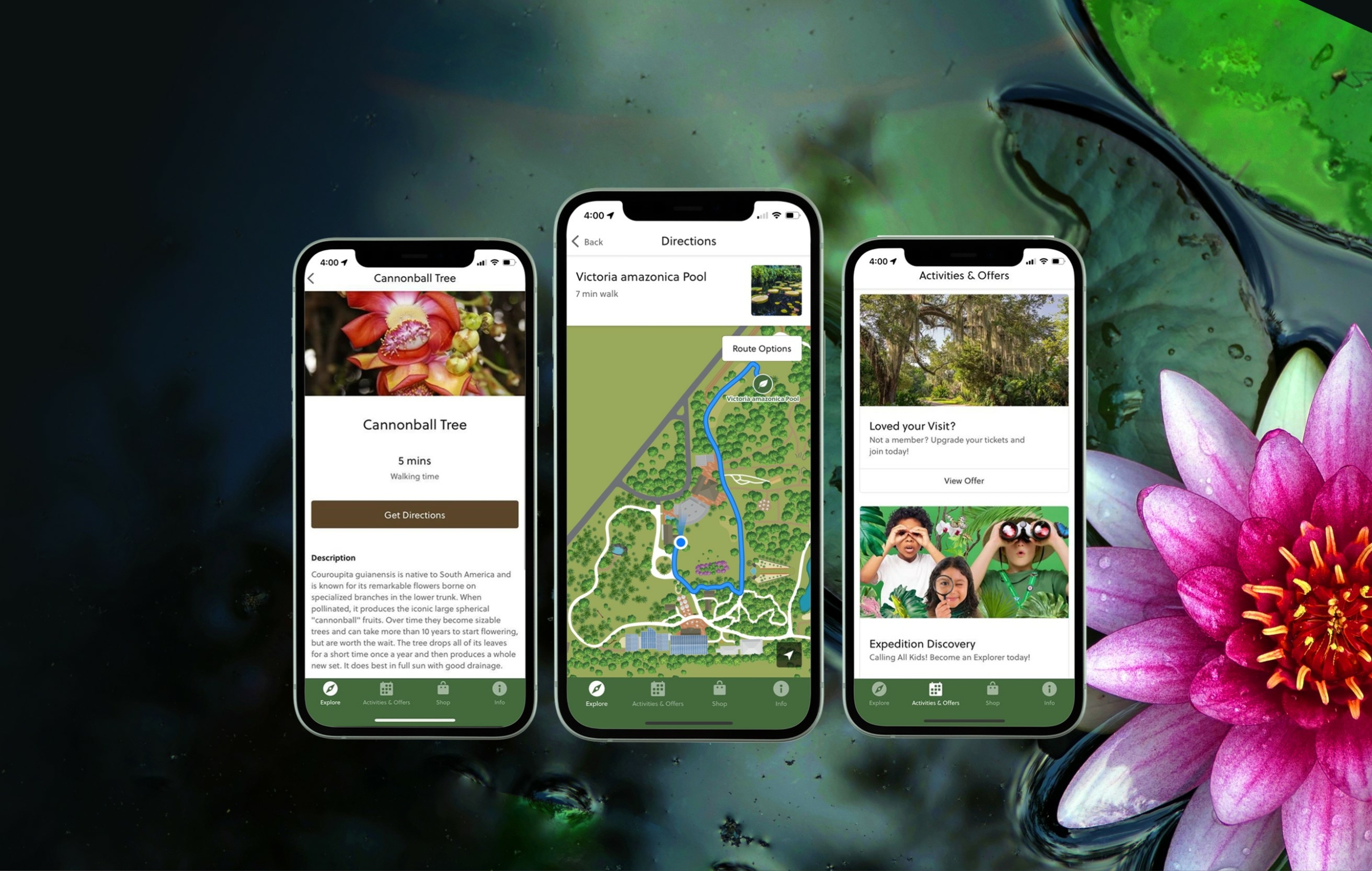



















Hours & Admission
Fairchild is open every day except for Christmas, December 25
Hours
10:00 a.m. – 5:00 p.m.
Admission
Fairchild Members Free
Adults $24.95
Child (6-17) $11.95
Students (with valid ID) $15.95
Seniors (65 and over) $17.95
In-Garden tram rides Free
Parking Free
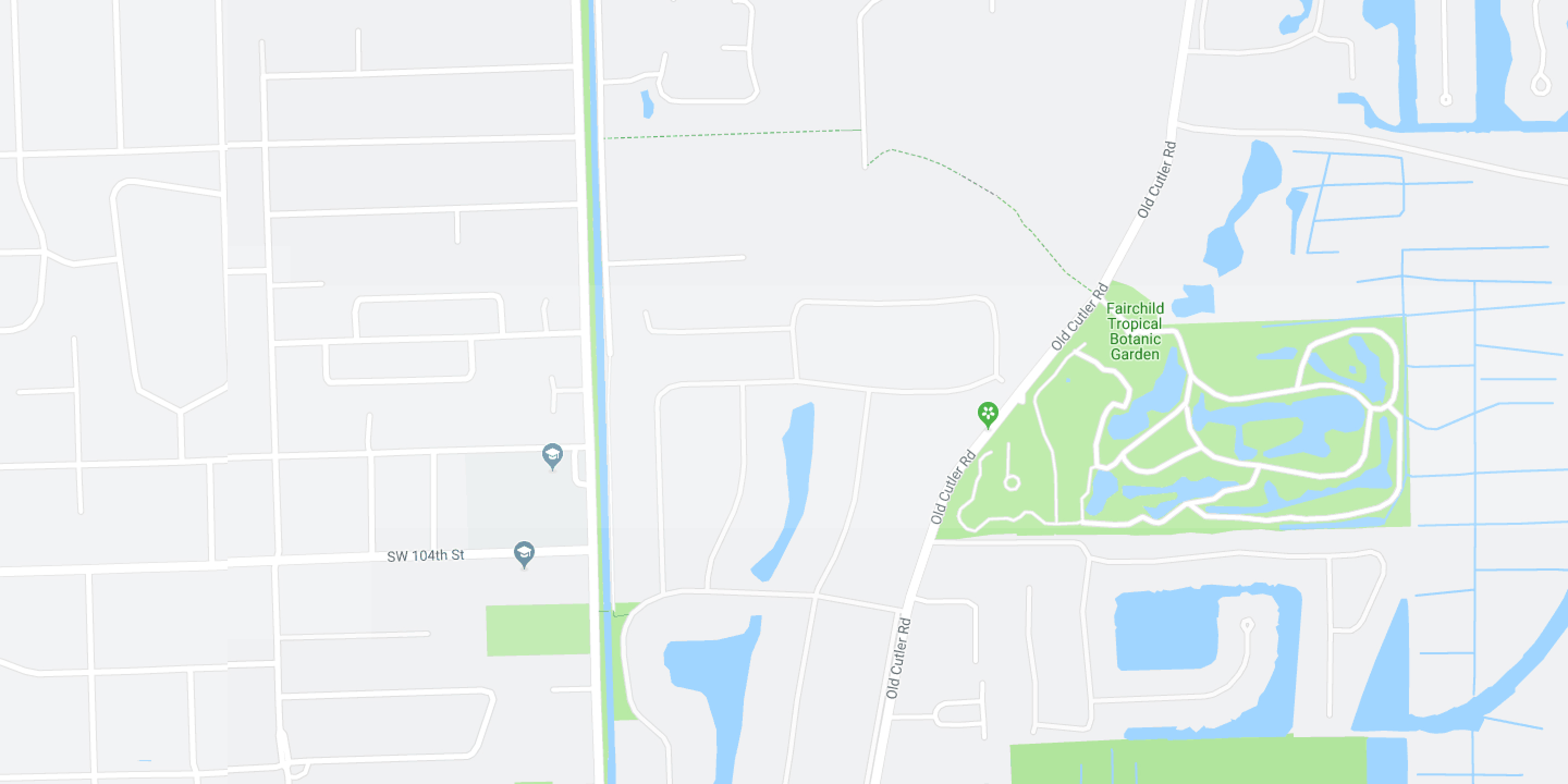
Location
10901 Old Cutler Rd, Miami, FL 33156
MembersGet Tickets Non-Members
Buy Tickets



FOLLOW THE LATEST @fairchildgarden